''ESTRUCTURA CELULAR''
En este capítulo se revisa la estructura y función básicas de los
componentes que constituyen a las células eucariotas y procariotas. El capítulo inicia con el análisis del microscopio. Desde el
punto de vista histórico, el microscopio reveló por primera vez
la presencia de bacterias y más tarde, los secretos de la estructura
celular. Hoy en día es aún una herramienta poderosa en el estudio de la biología celular.
MÉTODOS ÓPTICOS
El microscopio de luz.-
 El poder de resolución del microscopio de luz bajo condiciones ideales es de casi la mitad de la longitud de onda de la luz
utilizada. El poder de resolución es la distancia que debe separar dos puntos de fuentes de luz para que puedan observarse
como dos imágenes distintas.
El poder de resolución del microscopio de luz bajo condiciones ideales es de casi la mitad de la longitud de onda de la luz
utilizada. El poder de resolución es la distancia que debe separar dos puntos de fuentes de luz para que puedan observarse
como dos imágenes distintas.
En microbiología a menudo
se utilizan varios tipos de microscopios de luz:
a). Microscopio de campo brillante-. El microscopio de campo brillante es el utilizado más a menudo
en los cursos de microbiología y consiste en dos series de lentes
(objetivo y ocular) que actúan en conjunto para la resolución de
la imagen.
 b) Microscopio de contraste de fases.- El microscopio de contraste de fases se desarrolló para mejorar las diferencias de contraste entre las células y el medio circundante, con lo que se hace posible observar células vivas sin tinción; con los microscopios de campo brillante deben utilizarse preparaciones de microorganismos muertos y teñidos.
b) Microscopio de contraste de fases.- El microscopio de contraste de fases se desarrolló para mejorar las diferencias de contraste entre las células y el medio circundante, con lo que se hace posible observar células vivas sin tinción; con los microscopios de campo brillante deben utilizarse preparaciones de microorganismos muertos y teñidos.
c) Microscopio de campo oscuro.-
El microscopio de campo oscuro es el microscopio de luz en el
cual el sistema de iluminación se ha modifi cado para alcanzar
la muestra desde un solo lado. Esto se logra a través del uso de
un condensador especial que bloquea la luz directa y la refl eja a
través de un espejo ubicado a un costado del condensador en un
ángulo oblicuo
d) Microscopio de fluorescencia.-
 El microscopio de fluorescencia se utiliza para visualizar muestras con efecto de fluorescencia, que tiene la capacidad de
absorber luz de longitud de onda corta (ultravioleta) y emitir
luz con mayor longitud de onda (luz visible). Algunos microorganismos presentan fluorescencia natural por la presencia
de sustancias fluorescentes, por ejemplo la clorofila.
El microscopio de fluorescencia se utiliza para visualizar muestras con efecto de fluorescencia, que tiene la capacidad de
absorber luz de longitud de onda corta (ultravioleta) y emitir
luz con mayor longitud de onda (luz visible). Algunos microorganismos presentan fluorescencia natural por la presencia
de sustancias fluorescentes, por ejemplo la clorofila.
e) Microscopio diferencial de contraste
de interferencia (DIC).-
Los microscopios diferenciales de contraste de interferencia
uti lizan un polarizador para producir luz polarizada, la cual se
hace pasar a través de un prisma que genera dos haces distintos;
estos haces pasan a través de la muestra y entran al objetivo donde se combinan en un solo haz
ESTRUCTURA DE CÉLULAS EUCARIOTAS
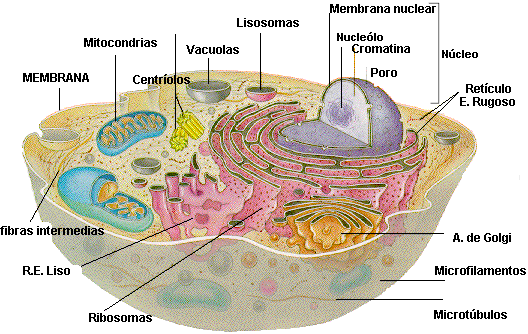
Núcleo.- El núcleo contiene el genoma de la célula. Está limitado por
una membrana formada por un par de unidades de membrana
separadas por un espacio de grosor variable. La membrana interna por lo común es un saco simple, pero la membrana más
externa se presenta en varios sitios como una continuación del
retículo endoplásmico
Estructuras citoplásmicas.-
El citoplasma de las células eucariotas se caracteriza por la presencia de un retículo endoplásmico, vacuolas, plástidos que se reproducen por sí mismos y un citoesqueleto complejo, compuesto por
microtúbulos, microfi lamentos y fi lamentos intermedios.
El retículo endoplásmico (ER, endoplasmic reticulum) es
una red de conductos limitados por membranas que tienen continuidad con la membrana del núcleo.
El aparato de Golgi consiste en un conjunto de
membranas que funcionan en combinación con el retículo endoplásmico para modifi car y organizar productos químicos del
retículo endoplásmico que más tarde serán secretados y aquellos
que participan en la producción de otras estructuras de la membrana celular.
Mitocondrias y cloroplastos. Varias pruebas sugieren que las mitocondrias y cloroplastos son
descendientes de microorganismos procariotas antiguos y que
se originaron del englobamiento de células procariotas por
células de mayor tamaño
Los lisosomas son sacos rodeados por membrana que contienen varias enzimas digestivas que las células utilizan para desdoblar macromoléculas como proteínas, grasas y polisacáridos.
El peroxisoma es una estructura rodeada por membrana
cuya función consiste en producir H2
O2
por la reducción de O2
a partir de varios hidrógenos donadores. El H2
O2
producido en
el peroxisoma más tarde se degrada a H2
O y O2
por acción de la
enzima catalasa.
El citoesqueleto es una estructura tridimensional que ocupa el citoplasma. Los tres tipos principales de fibras que comprenden el citoesqueleto son microfilamentos, filamentos intermedios y microtúbulos
Capas superficiales
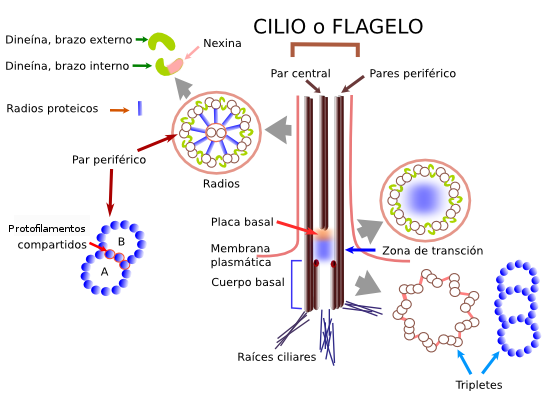
El citoplasma está rodeado por una membrana plasmática compuesta por proteínas y fosfolípidos, similar a la membrana de las células procariotas, ilustrada más adelante. La mayor parte de las células animales no tienen otras capas superfi ciales; no obstante, las células vegetales tienen una pared celular externa compuesta por celulosa.
ESTRUCTURA DE LAS CÉLULAS
PROCARIOTAS
La célula procariota es más simple que la eucariota al nivel de la
energía, con una excepción: la envoltura celular es más compleja.
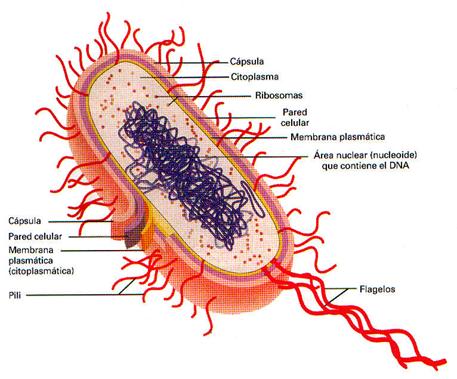
Nucleoide.- las células procariotas no tienen un núcleo verdadero; almacenan su DNA en una estructura conocida como nucleoide.
Estructuras citoplásmicas.- las células procariotas carecen de plástidos autónomos, como las mitocondrias y cloroplastos; las enzimas de transporte de electrones se localizan en la membrana citoplásmica.
Envoltura celular.- protegen al microorganismo de entornos ambientales hostiles, como osmolaridadextrema, químicos nocivos e incluso antibióticos.
Membrana celular.-
- A. Estructura: se diferencia de aquella de las células eucariotas por la ausencia de esteroles y la única excepción son los micoplasmas que incorporan esteroles, como el colesterol, en sus membranas cuando se cultiva en medios que contienen esteroles.
- B. Función: las principales funciones de la membrana citoplásmica son:
- Permeabilidad selectiva y transporte de solutos.
- Transporte de electrones y fosforilación oxidativa en especies aerobias.
- Excreción de exoenzimas hidrolíticas.
- Transporte de enzimas y moléculas que participan en la biosíntesis de DNA, polímeros de la pared celular y lípidos de la membran.
- Portar receptores y otras proteínas quimiotácticas y otros sistemas sensoriales de transducción.
Pared celular.- La pared celular bacteriana debe su resistencia a una capa compuesta de diversas sustancias conocidas como mureína, mucopéptidos o peptidoglucanos. La mayor parte de las bacterias se clasifican como grampositivas (se tiñen de color violeta) o gramnegativas (se tiñen de color rojo o rosa pálido) con base en su respuesta al procedimiento de tinción de Gram, la diferenciación entre estos dos grupos refleja diferencias fundamentales en sus envolturas celulares.
A. Capa de peptidoglucanos
Polímero complejo que consiste de tres partes: una estructura básica, compuesta de moléculas alternadas deN-acetilglucosamina y de ácido N-acetilmurámico y un grupo idéntico de enlaces peptídicos cruzados. Elácido diaminopimélico es un elemento singular en las paredes celulares bacterianas; nunca se encuentra en las paredes celulares de las arqueobacterias o de células eucariotas.
B. Componentes especiales de las paredes celulares de bacterias grampositivas
Contienen cantidades considerables de ácidos teicoico y teicurónico, los cuales pueden constituir hasta 50% del peso seco de la pared y 10% del peso seco de la totalidad de la célula; algunas paredes de bacterias grampositivas pueden contener moléculas de polisacáridos.
C. Componentes especiales de las paredes celulares de bacterias gramnegativas
Las paredes celulares de bacterias gramnegativas contienen tres componentes que se encuentran fuera de la capa de peptidoglucanos: lipoproteínas, membrana externa y lipopolisacáridos.
1. Membrana externa.- sirve para proteger a la célula de sustancias nocivas, como las sales biliares. Por su naturaleza lipídica, es de esperarse que la membrana externa excluya también a moléculas hidrofílicas.
2. Lipopolisacáridos (LPS).- las paredes celulares de bacterias gramnegativas consisten en un glucolípido complejo, denominado lípido A (consiste en unidades de disacárido de glucosamina fosforilada a la cual se unen varios ácidos grasos de cadena larga), el cual está unido a un polisacárido constituido por una porción central y series terminales de unidades repetidas.
3. Lipoproteínas.- las lipoproteínas son la proteína más abundante desde el punto de vista numérico en las células gramnegativas (casi 700 000 moléculas por célula). Su función es estabilizar la membrana externa y fijarla a la capa de peptidoglucano.
4. Espacio periplásmico.- espacio entre las membranas interna y externa, contiene la capa de peptidoglucano y una solución de proteínas que se comporta como un gel, representa casi 20 a 40% del volumen celular.
D. Pared celular de bacterias acidorresistentes
Algunas bacterias, entre las que sobresale el bacilo tuberculoso (M. tuberculosis) y bacterias relacionadas poseen paredes celulares que contienen grandes cantidades de ceras, que consisten de hidrocarbonos ramificados complejos (con longitudes de 70 a 90 carbonos) conocidos como ácidos micólicos. La estructura hidrófoba confiere a estas bacterias resistencia a muchos compuestos químicos como detergentes y ácidos fuertes; estos microorganismos se denominan acidorresistentes.
E. Pared celular de las arqueobacterias
Tienen pared celular rígida compuesta de polisacáridos o de un peptidoglucano conocido comoseudomureína. Las arqueobacterias que tienen una pared celular de seudomureína son grampositivas.
F. Capas superficiales cristalinas
Las bacterias, tanto grampositivas, gramnegativas y arqueobacterias, poseen una capa bidimensional de subunidades cristalinas con disposición en entramado formada por proteínas o glucoproteínas (capa S) como los componentes más externos de la envoltura celular. Las proteínas de la capa S son resistentes a las enzimas proteolíticas y a los agentes desnaturalizadores de proteínas; la función de la capa S es incierta pero probablemente sea protectora.
G. Enzimas que atacan la pared celular
Las enzimas que degradan la pared celular bacteriana también se encuentran en células que digieren la totalidad de la bacteria, por ejemplo protozoarios y células fagocíticas de animales superiores.
H. Crecimiento de la pared celular
Para la división celular es necesaria la síntesis de la pared celular; sin embargo, la incorporación de nuevo material de la pared celular varía con la forma de la bacteria.
I. Protoplastos, esferoplastos y formas L
La eliminación de la pared bacteriana puede lograrse con hidrólisis, con lisozimas o al bloquear la síntesis de peptidoglucano con un antibiótico como penicilinas; en los medios de cultivo con protección osmótica, tales tratamientos liberan protoplastos en las células bacterianas grampositivas y esferoplastos de las gramnegativas; si tales células son capaces de crecer y dividirse, se denominan formas L, y son difíciles de cultivar, por lo común requieren un medio de cultivo sólido con agar, además de encontrarse en un medio con la concentración osmótica adecuada.
J. Micoplasmas
Son bacterias que carecen de pared y que no contienen peptidoglucano. La diferencia entre las formas L y los micoplasmas es que es posible la síntesis de mureína, por lo que las formas L pueden adquirir nuevamente su forma original de bacterias, lo que nunca ocurre con los micoplasmas. Los micoplasmas carecen de un sitio de acción para los fármacos antimicrobianos que inhiben la síntesis de la pared celular (p. ej., penicilinas y cefalosporinas) y por tanto son resistentes a tales fármacos.
TINCIÓN
Los colorantes sufren combinación química con el protoplasma
de la bacteria; si la célula no está muerta, el proceso de tinción
la destruye; por tanto, tal proceso es drástico y puede producir
artefactos
Tinción de Gram-.

Una característica taxonómica importante de las bacterias es su
respuesta a la tinción de Gram. Las propiedades de tinción de
Gram parecen ser fundamentales, porque la reacción de Gram
se correlaciona con muchas otras propiedades morfológicas en
formas con relación fi logenética. Un microorganismo
que en potencia es positivo para la tinción de Gram puede parecerlo sólo bajo condiciones ambientales particulares y en un
cultivo joven.
Tinción acidorresistente.-
 Las bacterias acidorresistentes son aquellas que conservan la
carbolfucsina incluso cuando se decolora con ácido clorhídrico
en alcohol. Un frotis de células sobre una laminilla se cubre con
carbolfucsina y se calienta en baño María. A continuación se lleva a cabo la decoloración con la mezcla de ácido-alcohol y por
último se aplica una tinción de contraste.
Las bacterias acidorresistentes adquieren un color rojizo en tanto que
otras células adquieren el color del segundo colorante.
Las bacterias acidorresistentes son aquellas que conservan la
carbolfucsina incluso cuando se decolora con ácido clorhídrico
en alcohol. Un frotis de células sobre una laminilla se cubre con
carbolfucsina y se calienta en baño María. A continuación se lleva a cabo la decoloración con la mezcla de ácido-alcohol y por
último se aplica una tinción de contraste.
Las bacterias acidorresistentes adquieren un color rojizo en tanto que
otras células adquieren el color del segundo colorante. 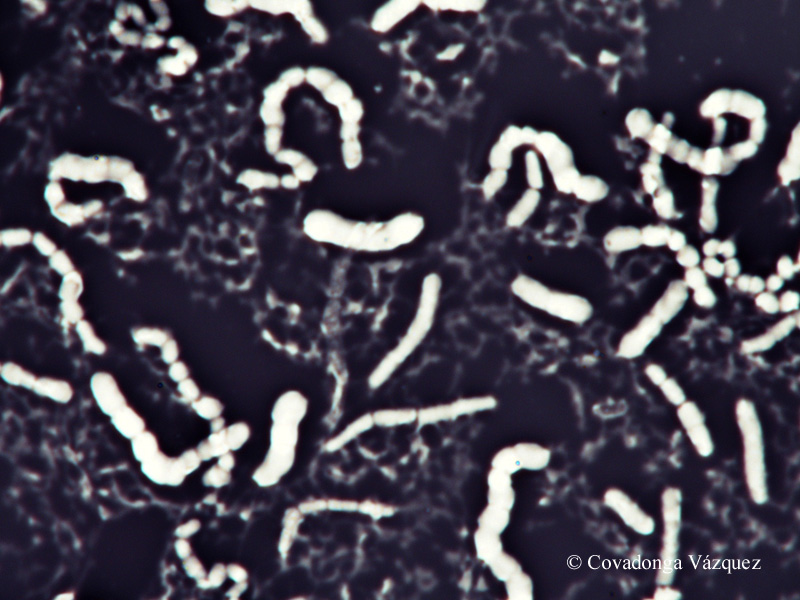
Tinción negativa.-
Este procedimiento consiste en la atención del entorno con un
colorante ácido, dejando a las células incoloras. El colorante negro de nigrosina (tinta china) se utiliza a menudo. Dicho método se emplea para aquellas células o estructuras difíciles de teñir
en forma directa
Tinción de los flagelos.-
 Los flagelos son demasiado delgados para que sean visibles en el microscopio de luz. Sin embargo, su
presencia y distribución pueden demostrarse al tratar las células
con una suspensión coloidal inestable de sales de ácido tánico,
lo que causa la precipitación intensa sobre las paredes celulares y
flagelos
Los flagelos son demasiado delgados para que sean visibles en el microscopio de luz. Sin embargo, su
presencia y distribución pueden demostrarse al tratar las células
con una suspensión coloidal inestable de sales de ácido tánico,
lo que causa la precipitación intensa sobre las paredes celulares y
flagelos Tinción de la cápsula.-
Tinción de la cápsula.-
La presencia de la cápsula por lo común se demuestra por procedimientos de tinción negativa o modifi caciones de tales procedimientos. Uno de estos métodos de “tinción de la cápsula” (método de Welch) implica el tratamiento con solución de
violeta de genciana caliente seguido de un lavado con solución
de sulfato de cobre

Tinción de esporas.-
Las esporas se observan de la manera más simple como cuerpos
refringentes intracelulares en suspensiones celulares no teñidas
o como áreas incoloras en células teñidas por métodos convencionales
No hay comentarios.:
Publicar un comentario